


马炜梁,植物学家。
不像动物,比如老虎要交尾的时候,两个雄虎拼命打,打到最后哪个胜了,哪个就获得了交配权。植物虽然不会动,但也有选择伴侣的能力,这是植物一个巧妙的地方。
植物开花的“智慧”
2021.12.12上海
各位好,我叫马炜梁。我是华东师范大学生命科学学院的退休教师,今年86岁。我从60年代开始工作,一直到1989年退休,到现在又过了三十多年。
我是做植物分类学的,平时上课就上植物分类学。我们做植物分类的应该算是做植物研究的人里认识植物最多的,做生态调查,就要请我们分类学的老师一起,因为碰到奇怪的植物,别人都不认识。因为要经常做野外考察,就去了很多地方。
下面这张照片是在海南岛拍的,当时刚从热带雨林考察完回来。热带雨林下雨是很猛的,我没带雨衣,就摘了一棵芭蕉的叶片当雨衣,把拍摄的器材放在前面用手捂着,就这样回到林场的驻地。

这是在四川的高山上,从这里的平地往上爬,开始是常绿阔叶林,后来是常绿落叶混交林,然后是矮林。在最高的地方,它后面已经没有树木了,山上也没有植物,这个地方就叫流石滩。

流石滩是植物分布的最高地带,这里岩石缝隙中的水结成冰以后体积增大,就把岩石崩开,崩开以后,岩石就沿着坡面落下来。
在流石滩下方的不远处,我看到一种植物,叫蚤缀,就拍了一张照片。这个植物像一个坐垫一样,半球形的,很奇特。它长成这样,风吹过来不容易倒伏,背风面在低温环境下不容易降低温度,更适应这里的寒冷气候。植物的垫状现象在高寒山地非常常见。

这是天山雪莲,只在天山有,它下部的叶片是绿的,上部是淡黄色半透明的苞片,就像一个温室的屋顶,保护花朵正常发育。等到花朵发育成熟,里面的花准备开放的时候,温室就像下面这张照片里一样打开了,以便昆虫进来传粉。

野外考察非常辛苦,但也有很多乐趣。我带同学们在野外实习的时候,啊呀,大家看到植物那么精彩,都很兴奋,和死气沉沉的课堂氛围真是天差地别。
为什会这样呢?因为当时植物分类学课本上的知识冷冰冰的。讲到一种植物,比如一棵树,只有几张彩色插图和一些概念,完全不能展示植物的活力,而且没有细微的解剖图片,我也没有办法把它的结构——雌蕊雄蕊、子房胚珠这些东西讲清楚,因此同学接收到的东西就比较空,上课提不起兴趣。
那么我就想,能不能把同学们对野外的兴趣引导到课堂上呢?
因为我初中的时候跟着父亲学过摄影,有摄影基础,所以从60年代开始就有个想法,能不能用什么镜头可以很清楚地把植物花、果的内部结构拍下来?我就到南京东路上的冠龙照相馆去问,它当时是上海最大的照相馆,也卖摄影器材,来来回回问了好多次,都说没有这样的镜头。直到80年代初的一天,我突然听朋友说现在有微距镜头了,就赶紧去买,花了3000多块买回来。
器材买来以后,我就试着做了两个植物的精细解剖,拍下来在课堂上面放,发现同学们简直太高兴了,特别喜欢。
后来我打电话去问国内一些比较老牌的学校,有没有植物精细解剖的照片,课堂上可以用,问了一圈都说没有。没有现成的材料,但同学们的课堂反馈又这么好,那怎么办呢?只能自己来,这个时候就下了这个决心,一定要把这件事情做下去。
这样我三十多年来,包括退休以后,就专门在家里面做植物的精细解剖。
这是我的工作台,最关键的是微距镜头,后面是一个解剖镜。先在解剖镜下解剖,然后再拿到微距镜头下拍,有了微距镜头以后,拍下来的东西就很清晰了。

这是马利筋。马利筋的果实散出来的时候,它的种子会粘在一起飘,有的时候你看到空中飘了一团棉花絮一样的东西,下面还挂着一个种子,那就是马利筋。

这是茅膏菜,它是吃虫的,我们叫它食虫植物。它的叶片上面全都是黏性的毛,如果昆虫经过这里,它就把昆虫的翅膀粘住,然后会慢慢地卷曲过来,把虫子消化掉。

这是刻叶紫堇和它的蒴果,图中间是我做了一个纵切。刻叶紫堇的种子成熟以后,它的两片果瓣会反卷,就像左边那样,把种子向各个方向弹出几十厘米。我为了拍照,就把它给固定住了,所以图中间的果瓣还没来得及把种子弹出去。

这是我自己用来做解剖的工具,都是很简单的器材。任意夹是我自制的,拍照的时候可以固定植物。夹东西的镊子要磨得很细很尖。刀片有单面刀片和双面刀片,而且双面刀片要把它的角去掉一点,变成个尖角,解剖的时候就很方便。

这是解剖开的香椿树的花,排列出来很好看。

这是棉花,棉花开花的时候做个纵切面,这样给学生们讲的时候就可以讲透了。
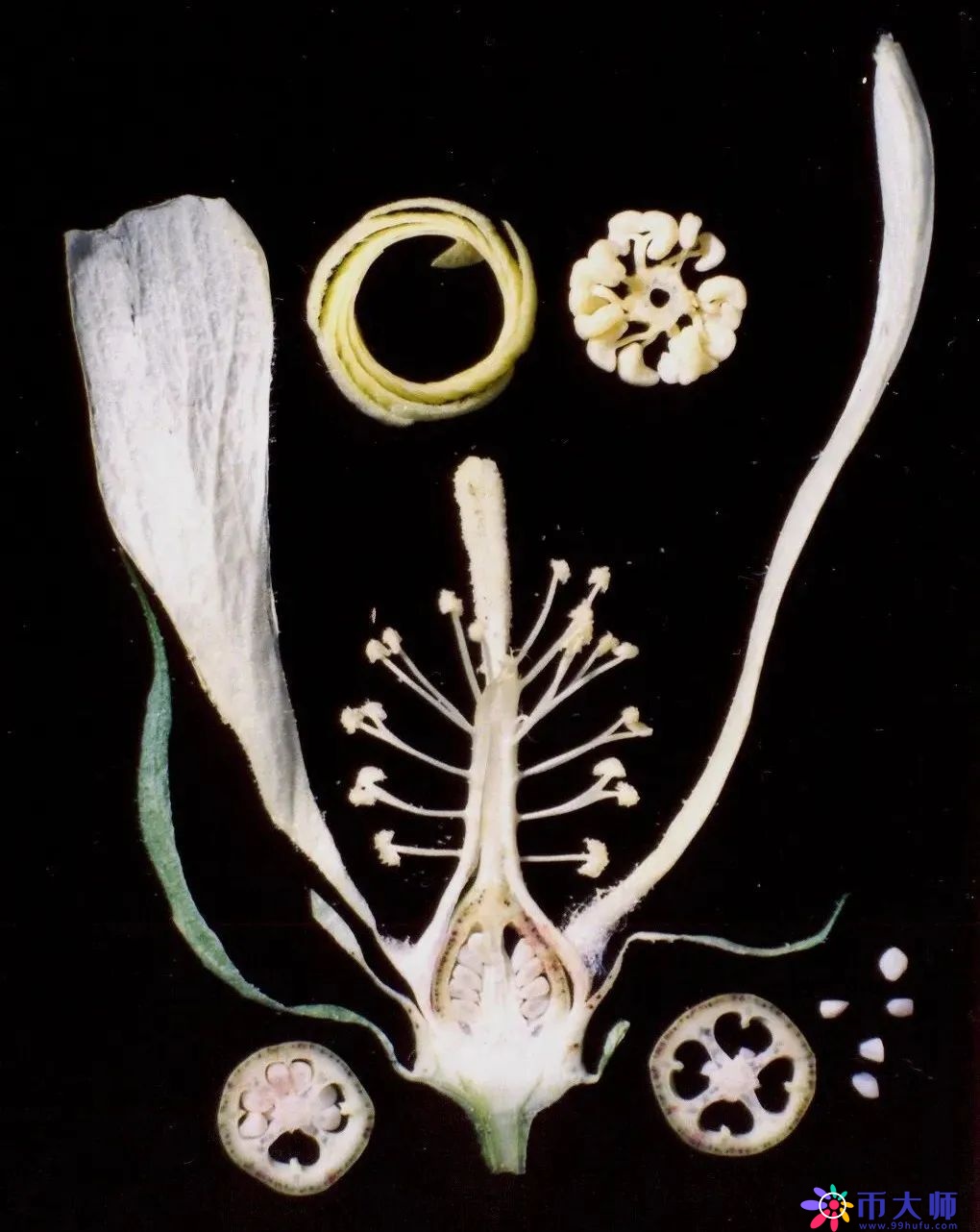
这是鹅掌楸的花,上面一圈是它的雄蕊,这么排列是因为它的雄蕊是螺旋状排列的。

这是上海的市花,白玉兰,也叫玉兰。这个花解剖开来蛮好看的,但实际上拍这些花的精细解剖图并不是为了好看,是为了教学。只有看得足够近,足够清楚,才能了解一朵花的内部结构。

接下来我就用我拍到的这些材料,为大家讲讲植物开花的“智慧”。
为什要讲开花?因为花朵对植物繁殖后代来说是最关键的,在植物不断的进化过程中,花朵是最集中体现植物“智慧”的器官。我们从蜡梅花讲起。

这是上海植物园的一个蜡梅园,四周都是开放的蜡梅花。

这是一棵树上正在开花的两朵蜡梅。

大家观察一下这两朵花的花心,右边一朵它的雄蕊是翻向外面的,左边的一朵,雄蕊都靠在一起,看上去好像是两朵不同的花。过去我们只能讲到这样的程度,因为最精细的没法解剖拍下来,所以同学看到蜡梅就是这个印象。
我们如果拍得再精细一点会怎么样呢?这是上面右边那朵蜡梅花心的解剖图,我把它做个切片,前面后面切掉后,可以清楚地看到雌蕊和雄蕊。雌蕊由花柱、柱头、子房三部分组成,雄蕊包括花药和花丝。这个时候,蜡梅的雄蕊都靠在边上,和柱头是分开的。

这是左边那朵花的切片,我们清楚地看到,蜡梅的花丝围拢了起来,花心明显和上面的花不一样。

这是怎么回事呢?
其实,这是蜡梅为了避免自花传粉进化出的一种很巧妙的传粉机制。
蜡梅的雌蕊和雄蕊并不是同时成熟的,开花初期,雌蕊先成熟,柱头接受蜜蜂从别的花上带来的花粉,完成受精。尽管自身的雄蕊很饱满,但是花药没有裂开,只是静静地等候在周围。
等到一两天之后,雄蕊的花丝发生了一个非常惊人的变化,花丝由向外弯曲变成直直地挺立起来,组成一道屏障,挡在柱头的外面,阻断了柱头和外面的交流,雌蕊不再接受外面的花粉。

接着,雄蕊成熟,花药裂开,向外散出花粉,等待蜜蜂把它的花药带给其他蜡梅。到这个时候,这朵花才算完成了开花的任务。
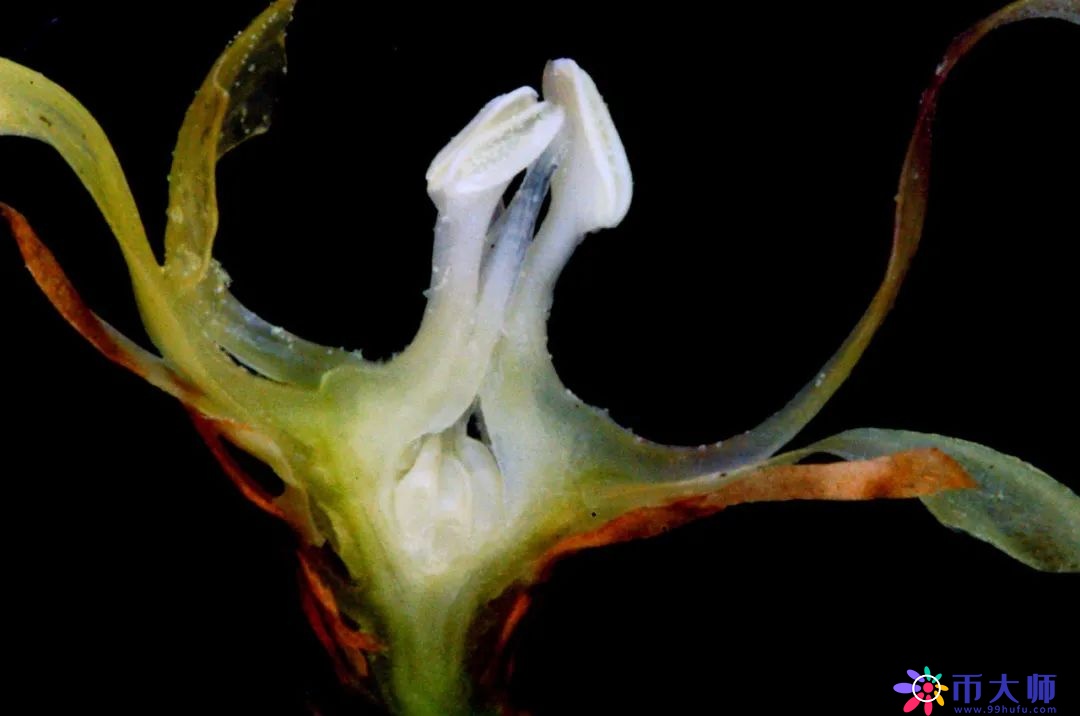
就是靠着雌雄蕊不同时成熟,以及立起的花丝的阻隔作用,避免了同一朵蜡梅花雌雄蕊的接触,多么巧妙。
我们再来看这张照片,考考大家,这两朵花哪一朵花先开放的?
这朵花是两性花,有雄蕊,有雌蕊,而且它们离得也很近,自己的雄蕊的花粉给雌蕊的柱头不是很方便吗?不行。为什么呢?大家知道,我们人跟动物一样,近亲繁殖会导致后代的衰落,植物也是这样的。
植物出现在地球上的时间远远早于人类,在不断地演化当中,自花传粉的植物后代在竞争当中逐渐被淘汰。那么植物在演化的实践当中就逐渐“懂得”了要避免自花传粉,利用在进化中“懂得”的这个道理,它们就演化出各种各样的传粉策略。
小小的蜡梅花通过雌雄蕊不同时成熟的传粉策略,自觉遵循着避免近亲繁殖的规律,而我们人类近200多年才通过现代遗传学解开了其中的原因。
不像动物,比如老虎要交尾的时候,两个雄虎拼命打,打到最后哪个胜了,哪个就获得了交配权。植物虽然不会动,但也有选择伴侣的能力,这是植物一个巧妙的地方。

一般来说,虫媒花的花朵开放以后,只是静静等待传粉昆虫的来访,传粉对植物来说是被动的,但也有例外。为了传粉,植物有时候也会采取一些主动行为。
这是日本小檗,在公园里很常见。我选了三个枝条,左边一个是开花的,当中一个是把它翻了过来,右边的一个已经结果实了。

小檗的花瓣有六片,它也是两性花,一朵花同时有雄蕊和雌蕊。在每朵花瓣的下部,有两块充满蜜汁的、深黄色的蜜腺。

当蜜蜂来采蜜时,必然用它的口器去接触蜜腺。蜜腺一被触碰,花瓣连带躺在它上面的雄蕊,一起向蜜蜂轻轻地扑打过去。

通过这么“啪”地拍打一下蜜蜂,小檗就把花粉撒到蜜蜂身上。
小檗的一朵花有6个花瓣,蜜蜂要是在每个花瓣的底部采一次蜜,可能“挨揍”6次,最后带着满身的花粉前往另一朵花传粉。
小檗科的植物大多具有拍打昆虫的行为,它们在全国分布很广,花期各异,是很好的观察材料。大家下次到公园时,可以用小檗枝条上的细刺模拟蜜蜂来访试试看。

构树这个植物也很有意思。有一次安徽钢铁厂的工人寄来一枝标本,说他们车间门口有一棵树会冒烟。这个实在奇怪,树怎么会冒烟呢?其实是构树在散粉。
构树在我们上海很普遍,有的时候家里面会自然而然地长出一棵树,就是构树,它的生命力很强。

构树是雌雄异株的,一棵树上要么全是雌株,要么全是雄株。下面左边的是雌株,右边是雄株。

放在微距镜头下看,雄株的花序上面长了很多雄花,每一朵雄花都有四个花药。右边是它的雌株,雌株上长满了长长的柱头。

四个花药挤在一起,每个花药都想往上长,结果四个花药碰在一起张不开来,等了一会儿以后,一个花药终于弹了出来,向外伸展,别的花药也跟着弹了出来。所以它开花的时候是很用劲地“啪”一下,使得它的花粉全部都散到空气中去。

就像一个抽烟的人吸了一口烟,用嘴巴轻轻地吐出一股白色的烟,在三四十公尺外也可以看到。
当时人们看到这个树会冒烟,觉得很不可思议,以为这个树是可以消灾的,有人去抱一抱树,有人去磕头,实际上这是很普遍的一个现象。
构树传粉是靠风力的,一棵树的花粉在空气中散出来以后,另一棵树的雌蕊怎么接触到这些花粉呢?
构树自然有自己的办法。它的雌花上长满了长长的柱头,它的长度是子房的25倍。而且柱头上还有黏性的毛,所以空气中飞过来的花粉就被这些长长的、黏性的柱头给截胡了,它就完成授粉了。


我们再来看看马兜铃是怎么传粉的。马兜铃在我们国家分布很广,大部分地区都有。
下面左边一张是它开的花,右边是它结的果。它的果实成熟以后,会裂开来,里面有很多种子,风一摇晃就像行进中的马脖子上戴的铃,“叮咚叮咚”响,风吹来这个种子就飘出来,到处散布。

马兜铃的花就像一个军乐队的大喇叭,上面有一个开口,底下是通向花心的喇叭管。解剖开喇叭管,我们看到,喇叭管里面长满了毛,而且都是倒向的毛。

在马兜铃开花的季节,常会看见喇叭口部有小蝇在飞舞。苍蝇是喜欢臭味的,正好马兜铃也有一种臭味,这个臭味对我们人来说很微弱,但是对一只苍蝇来说,味道是很浓的了。
小蝇跑到喇叭口上,闻到里面飘出的香味,接着就要往里面走,但是我们刚看到,喇叭口上面的毛都是向里的,所以它走到一半的时候,翅膀下面都粘上了毛,阻止它退回去。它退不回去,所以一定是进到里面。
我们再看花里面是什么样。下面这张图里,一只小蝇经过喇叭管以后豁然开朗,遇到一个大的空腔,再也没有刺毛限制它,于是就在里面欢快地寻找食物。就在这个时候,它把身上沾着的从另一朵花上带来的花粉抖落在花心六角形的凸起物上面,这是马兜铃的柱头,柱头下面是六个雄蕊。就这样,小蝇算是完成了给马兜铃传粉的任务。

解剖开这朵花时,一只小蝇正在花心进食。
当它吃饱了想出去的时候,却发现出不去了。因为刚才进来时是顺着毛的,现在要逆向地爬出去,是根本不可能的。它只好在里面继续转悠寻找出路。
而柱头接受了花粉之后,花心部分产生了两个变化:首先,在接受了小蝇带来的花粉后,柱头不久就萎缩了;另外,贴在柱头下方的花药成熟,开裂散出花粉。小蝇在里面乱转的时候,又沾上了这朵花的花粉。
就在花药成熟被小蝇沾上的同时,喇叭管内的毛慢慢萎缩,给小蝇让出了一条飞出的通道,被关了几个小时甚至一个晚上的小蝇终于重获自由。

马兜铃花解剖图。左边是花的纵切。中间和右边2列为花部的横切。左边一列的雄蕊尚未开裂,喇叭管内依然有毛刺;右边一列柱头萎缩,花药已开裂,喇叭管内毛已萎缩。
当小蝇重获自由飞出去以后,看到另一朵正在开放的马兜铃花,它又自投罗网,接受了马兜铃对它的“幽闭”,再把从刚才这朵花上沾上的花粉带往下一朵花授粉。
而刚才完成受精和散粉任务的花,已经不需要小蝇进来了,于是它就把花被盖在口部。

马兜铃和小蝇是地球历史上协同进化而来的一对伙伴。除了小蝇,没有其他昆虫为它传粉。也就是说,马兜铃离开了小蝇将会绝种,但小蝇没有了马兜铃,照样还能去吃其他的腐败有机物。
小蝇的传粉作用对马铃兜如此性命攸关,因此它才在进化中生成了这么巧妙的和小蝇的时空配合,令人不得不感叹,大自然千万年来的选择是人类无法复制的。而对于马兜铃花精巧的形态、结构,以及开花过程中时间、空间的基因调控,不是任何一名科学家现在能回答的问题。

我们前面讲了虫媒花、风媒花的传粉,那么在水里的植物怎么传粉的呢?有一次我走在松江县的田埂上,在边上的一条沟渠里,突然看到苦草上面漂了很多白色的粉末,但是没有被水浸湿。这些粉末不断从苦草下面升起,我想它肯定是苦草散放的东西。

这时候已经11月份了,水很凉了,但粉末不下沉还是第一次看到,好奇之下,我还是卷起裤管下到水里面,想挖几株带回去观察、拍摄。
当我把苦草放到浴缸里时,才发现它原来是雌雄异株的。左边是雄株,右边是雌株。

在雄株的基部,可以看到一个个的佛焰苞,里面长有几百朵圆球形的雄花。右边是做了一个纵切,里面的每个小圆点就是一朵未开放的雄花。
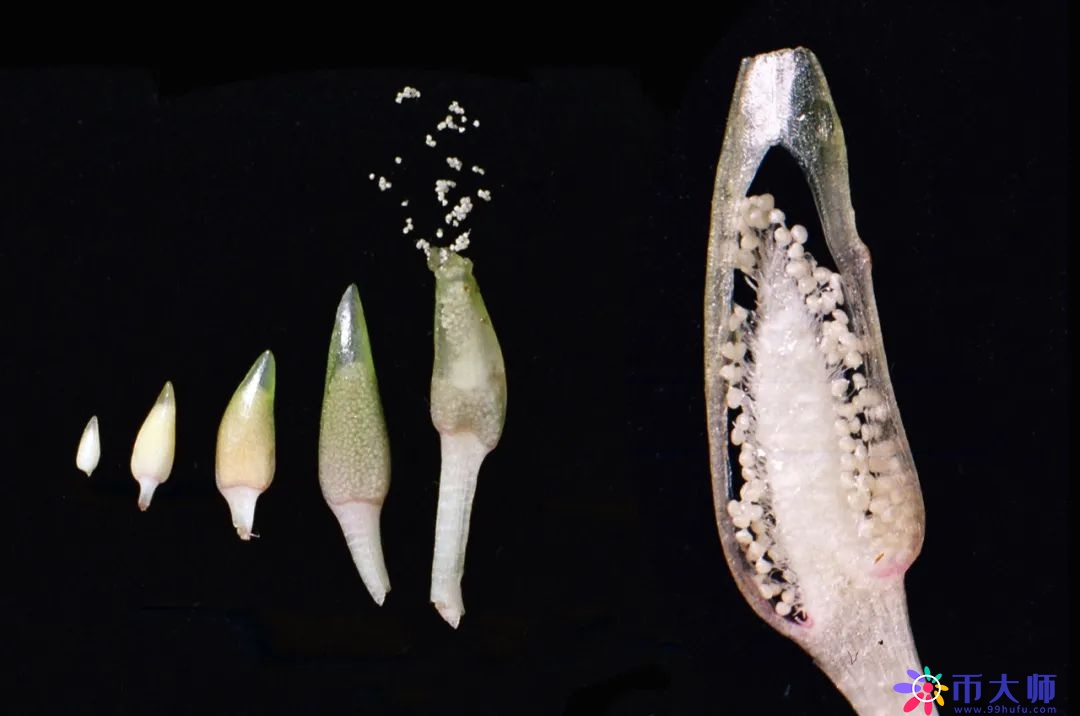
一旦成熟,佛焰苞顶上会开裂,里面的雄花就会一朵朵浮到水面上去,在静电作用下连成一片。之前在水面上看到的白色粉末就是它的雄花。
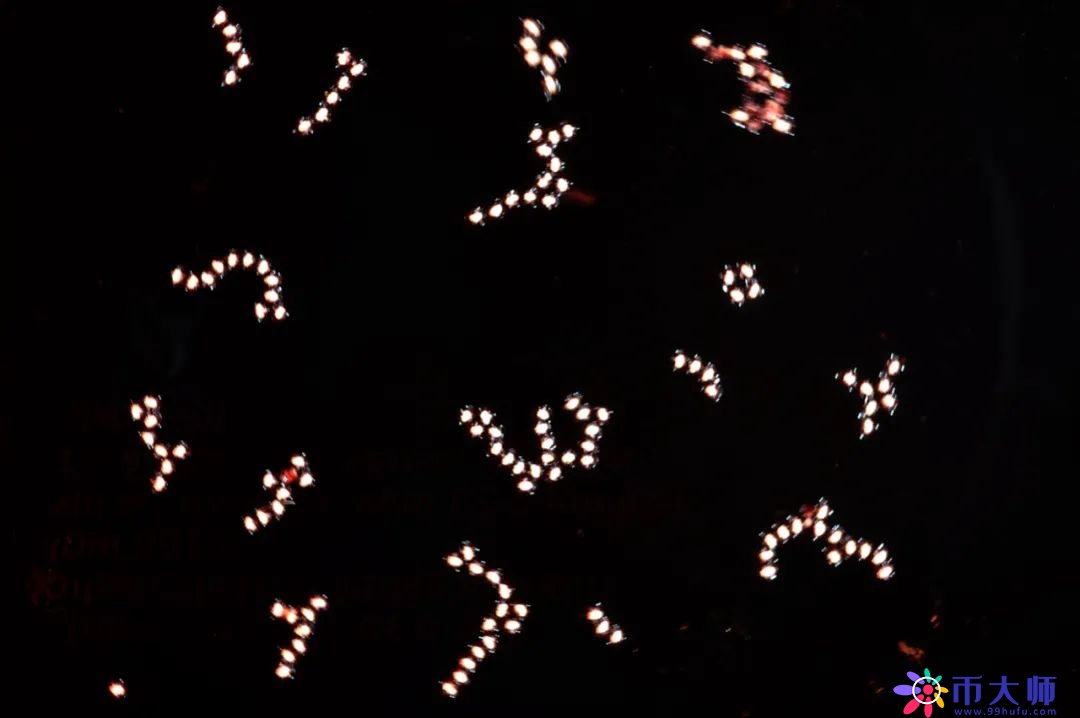
但是雄花浮到水面为什么不会被水打湿呢?放到显微镜下看,发现雄花升到水面以后,雄蕊周围的3片花被会张开来,像在花的边上架起3只小浮艇,利用浮力把花托举到水面上方1—1.5毫米的地方,使它不被打湿。因为花粉一旦被打湿,它就会沉到水底,影响它的传粉。
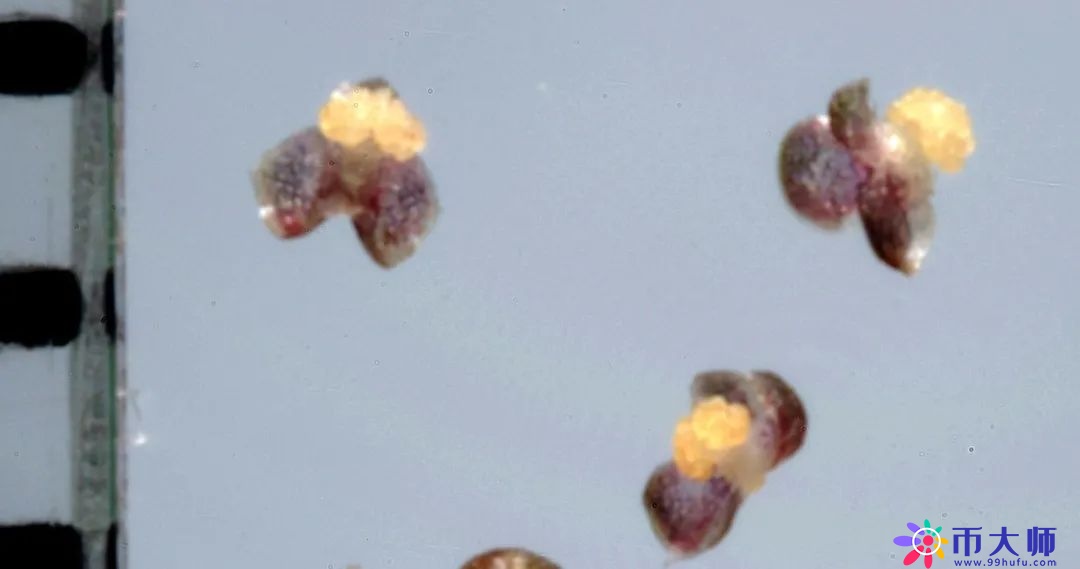
再看雌花,雌花的叶片是长条形的,两边有一个螺旋形的东西,像弹簧一样,是雌花的花柄。雌花要开放的时候,它的螺旋花柄就会把雌花托举到水面上开花。

雌花到水面之后,它柱头的位置刚好和雄花的花粉在同一水平面,就跟在水面上连成一片的雄花碰头了,受精就完成了。我是在茶杯的一角拍下这个照片的。

你看这个植物开花开得巧不,你雄花离水面1毫米,我雌花也1毫米,如果说你3毫米我只有1毫米,就碰不到了。
受精以后,雌花螺旋状的花柄会把雌花又拉入水下结果,使种子的发育得到母体充分的营养,等种子成熟以后,才撒手让它漂向远方。

水下生的苦草,这么巧妙地利用水的浮力完成了授粉,看了真是不禁拍案叫绝。

上世纪60年代,我从文献上看到,原产欧洲的无花果里边有一种小蜂专门为它传粉。我们都知道无花果看似无花,其实花都在隐头花序里,但隐头花序里面的花,靠什么昆虫来传粉呢?谁也说不清楚。
我们国家长无花果的地方还蛮多的,因此我每到一个地方,看到哪里的无花果结果了,都会采回来。但拿回来打开一看,里面都是花,根本找不到传粉的无花果小蜂,十几年的搜寻都没有结果。
1983年,我带学生到天目山进行野外实习。一位同学看到薜(bì)荔树上结的像馒头一样的果实——当地人叫鬼馒头,很好奇,就从树上采了一个,结果掰开一看,枯黄色的花里面全是虫,成千上百的虫,他吓了一跳,赶紧把它丢了。

采的时候我没看到,他丢的时候我看到了。我突然想到自己十几年寻无花果小蜂未遇的困惑,薜荔和无花果都是桑科榕属,都有隐头花序,薜荔里发现的这些虫,会不会解开我十几年来的困惑?于是赶紧把这个果实捡起来,放在包里面,带回驻地观察。
等回到驻地,重新打开这个果实,发现里面有两种颜色的小虫。我取出几朵枯黄色的花,放在解剖镜下准备拍照,发现有只黄虫蹲在花上,估计还没完全爬出来,于是请一位同学监视虫的动态,一旦爬出来,就可以拍下它的全貌。
大约20分钟后,这位同学突然高兴地叫起来:“马老师,出来了!”我跟他打趣说:“马老师一直在这这儿呢!”紧接着他又叫起来:“里面还有一只黑的!”

小蜂和它居住的花。黑色有翅膀的是雌蜂,黄色没有翅膀的是雄蜂。
其实这个发现是很值得一叫的,为什么呢?因为一朵花里装不下两只虫,那黄虫为什么趴在上面不下来,把尾部留在花内呢?它分明是在和黑虫交尾。若不是亲眼所见它们在交尾,你很难相信这两种体态判若两种的虫是同一种,我们把它叫昆虫的“二态现象”。
为了弄清楚这种虫的科学名称,我接连走访了6位昆虫学家。由于昆虫种类浩繁,这种小蜂不属于他们研究的目、科、属的任何一种,都遭到了谢绝。最后,浙江农业大学的徐教授在中山大学的帮助下鉴定出它的学名是薜荔榕小蜂。
那么,薜荔榕小蜂和薜荔是怎样的关系呢?为什么小蜂会在薜荔的花里交尾?
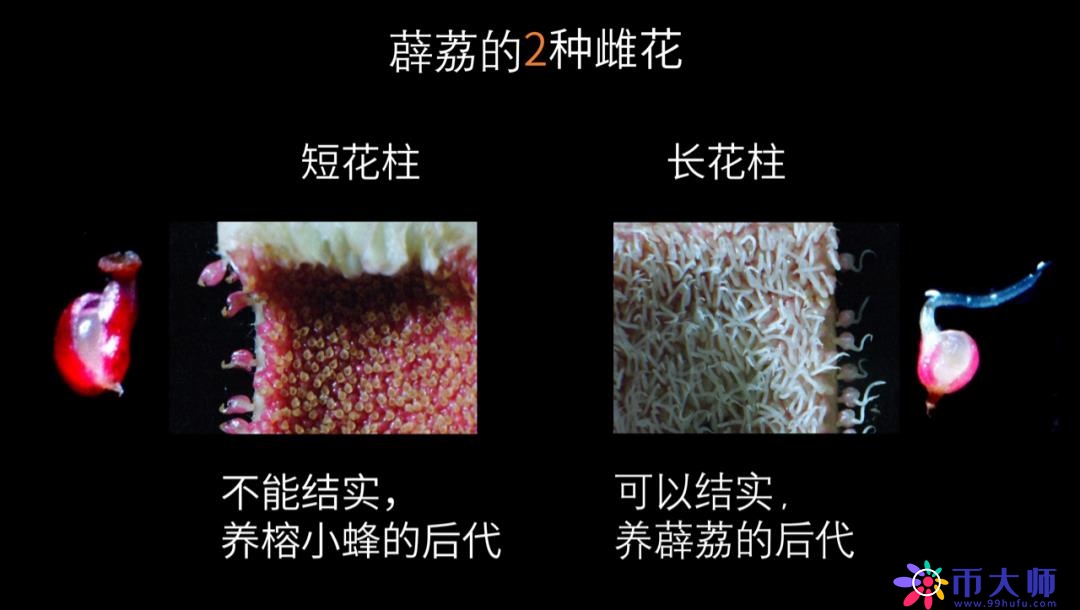
我们剖开一个薜荔的隐头花序看到,靠近开口部位的是1000多朵雄花,下面枯黄色的全是雌花。但很奇特的是,这种雌花并不是用来养育薜荔的后代的,而是用来养育榕小蜂的。

雌花的花柱,也就是连接柱头和子房的部分,它的长度刚好和榕小蜂的产卵器长度一样,榕小蜂把产卵器伸进去,正好到达子房。于是,小蜂就在雌花里发育,成熟交尾后,小蜂再外出寻找年幼的花序产卵。当它飞出时,会沾上花序上部雄花的花粉,顺便把它的花粉带往下一个花序。
那问题是,薜荔的后代谁来养育呢?
实际上,薜荔还有另一种花序,在这种花序里,只有雌花,但它的花柱比养育小蜂的那种雌花花柱更长,称为长花柱花。小蜂进入长花柱花后,它产卵器的长度到达不了子房,找不到可以产卵的地方,它到处兜,在到处兜的时候,它翅膀上的花粉都落下来,掉到长花柱花的柱头上,起到了传粉的作用。
长花柱花受粉以后就会长出薜荔的果实。而误入长花柱花的小蜂耗尽体力后,满腹怀卵,死在花序内。
那么,为什么薜荔要养育小蜂的孩子,而小蜂为什么又会“自杀式”进入薜荔的花序,这对它们有什么好处呢?
实际上,在漫长的进化当中,薜荔和榕小蜂相互适应,协同进化,最后形成了专一性共生的关系,这是自然界动植物之间相互关系的最高形式。薜荔和薜荔榕小蜂,其中一方离开另一方,将会绝种。
短花柱花序的薜荔像哺育自己的胚胎一样为榕小蜂的幼虫提供营养,它整年的光合作用产物几乎全花在培养几千只小蜂上面了。看起来薜荔做出了巨大的牺牲,但养育的榕小蜂的后代可以为薜荔传粉,使得薜荔的后代发展壮大。
而榕小蜂“自杀”式飞入薜荔的长花柱花序,为薜荔传粉,似乎也是一种无偿的牺牲,其实也不然。由于它的传粉,雌花序中结出了大量的种子,当这些种子若干年后长成了大树,榕小蜂的后代何愁找不到更多的栖息与繁育场所。

这是很奇妙的,它的结果就是使得薜荔和小蜂都得到繁衍。
我们前面讲到植物开花的“智慧”,其实这只是形象的说法,看起来蜡梅、日本小檗、马兜铃、苦草、薜荔开花好像很有智慧的样子,但其实这只是植物在漫长的进化当中由大自然随机、无目的地选择的结果。

我上面所讲的内容都来自我做植物精细解剖的材料。我前面讲下定决心自己来拍植物的精细解剖,话虽这么说,但植物分类学的课程涉及的植物种类众多,这些植物的生长环境各异、开花季节不同,如何能完成这一任务呢?这必然是一场旷日持久的工作。因此拍着拍着,我又发动了其他几位同志来一起完成。
为了得到新鲜的花、果,必须人随花动,花在什么季节开,在什么地方开,就应该此时到达此地,否则必然错过了最好的拍摄时机,而两种生长在不同地域的植物在同一季节开花了, 没有“分身术”的我们又只能等到下一年了。

就这样见缝插针地做,每做一种就积累一种,经常在外出考察的时候利用本职工作的间隙拍摄到半夜,30年积累下来,最终拍完了1008属植物,照片拍了7500多张,最后编成《中国植物精细解剖》一书出版。回过头来看,30年前虽然下定了决心,但能完成这件事在当时还是不可想象的。
因为我退休了,拍摄的160多个胶卷放在系里也没人看管,最后就送给中国科学院植物研究所,他们拿到恒温室去保存。
除了《中国植物精细解剖》,我还出了一本科普书,叫《植物的“智慧”》,这是针对学生在学习中的问题而写的。各位如果对植物的知识感兴趣,可以去读这本书。

这两本书是我30年工作中的副产品。单位不是要我退休嘛,我退休了,但退休了只是职业结束了,我的事业并没有结束,所以我在家里面做精细解剖,做了30多年。
谢谢各位,我就讲到这里。
本站所有软件信息均由用户上传发布,版权归原著所有。如有侵权/违规内容,敬请来信告知邮箱:liqiaoqiant@qq.com,我们将及时撤销! 转载请注明出处:https://www.99hufu.com/baike/204490.html